Bluetongue Virus
Bluetongue virus (BTV), a member of Orbivirus genus within the Reoviridae family causes serious disease in livestock (sheep, goat, cattle). Partly due to this BTV has been in the forefront of molecular studies for last three decades and now represents one of the best understood viruses at the molecular and structural levels. BTV, like the other members of the family is a complex non-enveloped virus with seven structural proteins and a RNA genome consisting of 10 double-stranded (ds) RNA segments of different sizes. Data obtained from studies over a number of years have defined the key players in BTV entry, replication, assembly and exit and have increasingly found roles for host proteins at each stage. Specifically, it has been possible to determine the complex nature of the virion through 3D structure reconstructions (diameter ~ 800 Å); the atomic structure of proteins and the internal capsid (~ 700 Å, the first large highly complex structure ever solved); the definition of the virus encoded enzymes required for RNA replication; the ordered assembly of the capsid shell and the protein sequestration required for it; and the role of host proteins in virus entry and virus release. These areas are important for BTV replication but they also indicate the pathways that may be used by related viruses, which include viruses that are pathogenic to man and animals, thus providing the basis for developing strategies for intervention or prevention.
BTV is the type species of the genus Orbivirus within the family Reoviridae. The Reoviridae family is one of the largest families of viruses and includes major human pathogens (e.g., rotavirus) as well as other vertebrate, plant and insect pathogens. Like the other members of the family, Orbiviruses which encompass, besides BTV, the agents causing African horse sickness (AHSV) and epizootic hemorrhagic disease of deer (EHDV), have the characteristic double-stranded and segmented features of their RNA genomes. However, unlike the mammalian reoviruses, Orbiviruses comprising 14 serogroups, are vectored to a variety of vertebrates by arthropod species (e.g., gnats, mosquitoes and ticks) and replicate in both hosts. BTV, the etiological agent of Bluetongue disease of animals, is transmitted by Culicoides species. In sheep BTV causes an acute disease with high morbidity and mortality. BTV also infects goats, cattle and other domestic animals as well as wild ruminants (e.g., blesbuck, white-tailed deer, elk, pronghorn antelope, etc.). The disease was first described in the late 18th century and was believed for many decades to be confined to Africa. However, to date BTV has been isolated in many tropical, subtropical and temperate zones and 24 serotypes have been identified from different parts of the world. Due to its economic significance BTV has been the subject of extensive molecular, genetic and structural studies. As a consequence it now represents one of the best characterised viruses.
Unlike the reovirus and rotavirus particles, the mature BTV particle is relatively fragile and the infectivity of BTV is lost easily in mildly acidic conditions. BTV virions (550S) are architecturally complex structures composed of 7 discrete proteins that are organised into two concentric shells, the outer and inner capsids, and a genome of 10 dsRNA segments. The outer capsid, which is composed of two major structural proteins (VP2 and VP5), is involved in cell attachment and virus penetration during the initial stages of infection. Shortly after infection, BTV is uncoated, i.e. VP2 and VP5 are removed, to yield a transcriptionally active 470S core particle which is composed of two major proteins VP7 and VP3, and the three minor proteins VP1, VP4 and VP6 in addition to the dsRNA genome. There is no evidence that any trace of the outer capsid remains associated with these cores, as has been described for reovirus. The cores may be further uncoated to form 390S subcore particles that lack VP7, also in contrast to reovirus. Subviral particles are probably akin to cores derived in vitro from virions by physical or proteolytic treatments that remove the outer capsid and causes activation of the BTV transcriptase. In addition to the seven structural proteins, three non-structural (NS) proteins, NS1, NS2, NS3 (and a related NS3A) are synthesised in BTV-infected cells. Of these, NS3/NS3A is involved in the egress of the progeny virus. The two remaining non-structural proteins, NS1 and NS2, are produced at high levels in the cytoplasm and are believed to be involved in virus replication, assembly and morphogenesis.
Structurally Designed Vaccines for BTV
Bluetongue disease affects mainly sheep and cattle and is economically an important disease in many parts of the world, both in developed and developing countries. Since it is a vector-borne disease and transmitted to healthy livestock from infected animals by blood feeding Culicoides spp., attenuated, live virus vaccines are not desirable. The flexibility of baculovirus expression vectors and the capacity of the baculovirus genome to accommodate large amounts of foreign DNA allowed us to exploit the system for the simultaneous expression of multiple BTV genes by a single recombinant virus. Based on the 3D structural data, recombinant viruses were prepared to express either VP3 and VP7, the two major core proteins, or VP3 and VP7 together with VP2 and VP5, i.e. all four major structural proteins. Expression of VP3 and VP7 resulted in CLPs that were similar in size and appearance to cores prepared from BT virions. Similarly, the simultaneous expression of four proteins resulted in the assembly of virtually homogeneous doublecapsid particles. When the 3D structure of CLPs and VLPs were analysed by Cryo-EM both types of particles were clearly comparable to authentic cores and virions, and exhibited essentially the same basic features and full complement of the two or four proteins. VLPs synthesized by recombinant baculoviruses were also characterized further at the biological and immunological levels and compared to those of the native virion. VLPs exhibited high levels of haemagglutination activity similar to those of authentic BTV. Further antibodies raised to the expressed particles contained high titres of neutralizing activity against the homologous BTV serotype emphasizing their authenticity at functional level.Since VLPs elicited strong neutralizing antibodies in guinea pigs, it can be anticipated that VLPs should also elicit protective responses in sheep against BTV infection. Consequently, a number of experiments were performed to examine the protective efficacy of VLPs (10-200 μg per sheep) in 1-year old BTV free sheep divided in groups (4-8 sheep/group). All sheep were challenged by subcutaneous inoculation of 1 ml of infective sheep blood containing virulent virus at day 117 or at 14 months. The challenged sheep developed neither clinical signs nor viraemia, indicating a suppression of BTV replication. The post-challenge blood samples of the sheep that did not receive vaccine and only received saline, contained infectious BTV and these sheep developed high neutralizing antibody titres indicative of a primary infection. Protective immunity to BTV disease was obtained by vaccinating sheep with doses of 10 μg or more of BTV VLPs which resulted in long-lasting protection against homologous BTV challenge. Some preliminary evidence was obtained for crossprotection, depending on the challenge virus and the amounts of antigen used for vaccination.
Future Perspectives
Bluetongue disease emerged in Europe only recently but is becoming one of the major threats to livestock industry world-wide. Although our knowledge of host-virus interactions and viral pathogenesis is limited, BTV is well characterised both genetically (the sequence was completed in 1989) and structurally. Understanding of molecular biology of the virus and mapping the role of each protein in virus life cycle has been benefitted significantly from the availability of recombinant BTV proteins and sub-viral particles. In addition the structure of BTV proteins, core and virion particles have contributed greatly to understanding the mechanism of protein-protein interaction in the virus assembly pathway of BTV and other orbiviruses. Most importantly, information gained from these studies has laid sound foundation for the generation of safe BTV vaccines with the possibility of use in animals in the near future. Latterly, studies have concentrated on the fundamental mechanisms that are used by the virus to invade, replicate in and escape from susceptible host cells. Progress has been made in understanding the structure and entry of intact virus particles, the role of each enzymatic protein in the transcription complex, the critical interactions that occur between the viral nonstructural proteins and viral RNA and the role of cellular proteins in non-enveloped virus egress.Despite these advances, some critical questions remain unanswered for the BTV life cycle and a more complete understanding of the interactions between the virus and the host cell are required for these to be addressed. For example, although progress has been made in the identification of signals for the recruitment of BTV RNA segments into the virion assembly site in the host cell cytoplasm, it has not been possible yet to determine exactly how each genome segment is packaged into the progeny virus. It is also not apparent when and how these genome segments wrap around the polymerase complex once the RNA has been recruited. Very little is known about the intracellular trafficking of newly generated virions although there are some indications of involvement of the cytoskeleton, intermediate filaments and vimentin during BTV morphogenesis. Host-virus interactions during virus trafficking will be one of the future areas needing intense attention. Recent work has revealed unexpected and striking parallels between the entry and release pathways of BTV and pathways involved in entry and release of enveloped viruses. These parallels may be the result of an enveloped ancestor virus; or because there are a limited number of cellular pathways that can be useful for the egress of large protein complexes from cells. It is notable that the NS3 glycoprotein of BTV is an integral membrane protein that is functionally involved in virus egress by bridging between the outer capsid protein VP2 and the cellular export machinery. Although no cell-free enveloped form of BTV has been isolated, budding of BTV particles from infected cells at the plasma membrane are quite apparent. The exact role of NS3 in this process and the role of host proteins (Annexin II and p11, Tsg101 and MVB) in the release of non-enveloped viruses, such as BTV, remain to be clarified.
One of the major drawbacks of research with BTV and other members of the Reoviridae has been the lack of a suitable reverse genetics system which allows genetic manipulation of the virus. This has been a major obstacle in understanding the replication processes of these viruses. However, recently it has been possible to recover infectious BTV wholly from ssRNA, which suggests a means for establishing helper virus-independent reverse genetics systems for members of the Reoviridae. The discovery that BTV ssRNA is infectious brings the realisation of a reverse genetics system for orbiviruses a step closer. There is no doubt that this will be one of the future dominant research areas for the members of the Reoviridae. Once in place, molecular and structural studies of individual BTV proteins can be placed in the context of the whole virus.
BTV outer capsid proteins, structural organization and their involvement in cell entry
The non-glycosylated outer capsid proteins of BTV are involved in virus entry to the host cells. A combination of three-dimensional (3D) structural determination and molecular/ biochemical analysis of the BTV outer capsid has given some insight into how the two outer capsid proteins could function in cell entry/membrane penetration processes in an analogous manner to enveloped virus proteins. Cryo-electron microscopy (Cryo-EM) and image reconstruction of BTV particles have been used to deduce the general relationships of the two outer capsid proteins within the virus particle. An initial low-resolution (45 Å) 3D reconstruction followed by a higher resolution (22 Å) reconstruction of BTV particles revealed a well-ordered morphology of the virion particle with icosahedral symmetry and provided details of the essential organization of the two outer capsid proteins. An accurate position of the two proteins, VP2 and VP5, in relation to each other and to the underlying protein layer of the core was clearly defined .
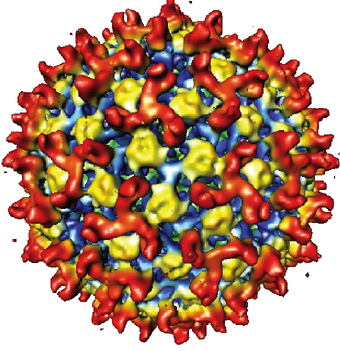
In addition, their juxtaposition and structural features gave an insight into their role in virus attachment to the host cell receptor and membrane penetration that are supported further by biochemical data. The outer capsid of BTV has an icosahedral configuration, and exhibits a unique organization made up of two distinct shapes, triskelion and globular. The outer layer does not conform to a T = 13 icosahedral symmetry like rotaviruses and reoviruses and consists of a total of sixty triskelion densities and 120 globular densities.
VP2, its structure and function
The most external part of the outer capsid is the propeller-shaped triskelion motif. At the tip of each blade of the propeller, the molecule broadens out and bends upwards perpendicular to the plane of the virus. These bent tips give the entire virion a diameter of ~880 Å and extend from the main body of the particle by 4 nm. The estimated volume of each triskelion structure closely matches that of three VP2 molecules. In solution recombinant VP2 oligomerises into a trimer readily, supporting the structural data. The structural features of VP2 are of a sailed-shaped, extended propeller blade on the virion surface and correlate well with the functional features of VP2. It is the most variable of the BTV proteins, eliciting virus neutralizing antibody, and is responsible both for serotype-specificity and virus haemagglutination activity indicating that VP2 is the cellular receptor binding protein. Direct evidence that VP2 indeed mediates attachment of the virion to the cell surface and its subsequent internalization has been obtained subsequently by biochemical studies using virion particles, recombinant particles and soluble VP2.
VP5, its structure and function
Interspersed between the VP2 triskelions and lying more internally are the VP5 globular densities that are also entirely exposed in the virion. Three molecules of VP5 (~60 kDa) conform to the globular density observed and as recombinant VP5 in solution also forms trimers, the 120 globular densities per virus particle comprise a total of 360 VP5 molecules. At the sequence level VP5 is divided into an amino terminal coiled-coil domain and a carboxyl terminal globular domain with a flexible hinge region in between. The N-terminus of VP5 also includes a membrane-inserting hydrophobic region followed by a long heptad repeat region analogous to the other fusion proteins. These features, as well as its location, support its involvement in membrane permeabilizing activity. In the absence of other BTV proteins, VP5 is highly cytotoxic and purified VP5 protein was shown to have intrinsic membrane 'pore-formation' ability, an activity that maps to the amphipathic helices.
That VP5 is indeed the BTV membrane penetration protein has been formally demonstrated by an experimental system where VP5 was expressed on the cell-surface. By this system it was possible to show that VP5 has the ability to interact with host cell membranes and to induce cell-cell fusion in a manner similar to that seen for the fusion proteins of enveloped viruses. VP5 exhibits its membrane-interacting properties only after it has undergone a low-pH triggered activation step, which presumably alters the conformation to fusion-competence. Deletion of the first 40 residues (the two predicted amphipathic helices) abolished fusion activity and, notwithstanding the precise mechanism, it is clear that VP5 can functionally substitute for a typical viral fusion protein. Although it is not yet clear if fusion induced by VP5 follows the same mechanisms proposed for enveloped viruses, it seems probable that the amphipathic helices insert into the target membrane. This early interaction and the formation of the initial pore might be mechanistically different for VP5 when compared with other fusion proteins as the amphipathic helices of VP5 more closely resemble pore-forming antimicrobial peptides than typical fusion peptides. Thin section EM studies exhibited the presence of BTV particles in coated pits, suggesting clathrin-mediated endocytosis may be used for BTV entry. A recent comprehensive investigation using biochemical and confocal microscopy studies in combinations of various specific inhibitors and RNA interference conclusively demonstrated that indeed BTV enters cells by clathrin-mediated endocytosis and requires acidic pH-dependent penetration.

Notably, in the BTV particle the VP5 protein makes strong contact with the underlying VP7 layer and also with the spike protein VP2 which protrudes above VP5 and has direct contacts with underlying VP7 as well. Thus, a plausible hypothesis would be that the attachment of VP2 with receptor and subsequent low pH inside endosomes induce a rearrangement and conformational changes in VP2 and VP5 which result in loosening the interactions of VP5 and VP7 and simultaneously allowing VP5 to form a protein layer with intrinsic outside-in curvature. Proteolytic removal or pH induced conformational change of VP2 then allows the VP5 amphipathic helices to freely interact with membranes and initiate the permeabilization process. This allows core particles that now lack both VP2 and VP5 proteins, to be released into the cytoplasm from the endosomal vesicles.
Further reading
- Foot-and-Mouth Disease Virus: Current Research and Emerging Trends
- Influenza: Current Research
- Virus Evolution: Current Research and Future Directions
- Arboviruses: Molecular Biology, Evolution and Control
- Alphaviruses: Current Biology